细菌信号转导网络复杂度的进化原理获得新认知
中国科学院南海海洋研究所热带海洋生物资源与生态重点实验室研究员高贝乐团队以具有足够进化深度和生态多样性的弯曲菌门为研究对象,分析了六大信号转导系统在该门不同分支的进化过程及复杂度构建的方式,解析了细菌的信号网络从简单演变为复杂,或从复杂至简单的过程。相关研究成果近日以“Evolutionary Principles of Bacterial Signaling Capacity and Complexity”为题发表于mBio。
细菌依赖信号转导系统来感知和响应环境变化以维持生存和繁殖,目前已知的细菌信号转导系统主要有六类:1.趋化系统,主要指挥细菌的运动方向; 2.双组份信号转导系统,调节基因的转录与表达;3.第二信使介导的信号转导系统,例如环化腺苷酸、环二鸟苷酸;4.STYK磷酸激酶和磷酸酶介导的信号通路;5.ECF介导的信号通路,调节基因的转录和表达;6.用于细菌间交流的群体感应。这些信号转导系统可以独立存在,也可以交互在一起,形成一个网络。以往的调查研究对不同细菌物种中的几个信号转导系统的组成蛋白进行了统计和分类,以了解它们的系统组成和多样性、模块化和可塑性、系统发育分布、与基因组大小和生态分布的相关性等等。然而,目前尚未完全清楚细菌的信号转导网络在整体上是如何演变的。
回答该问题,需要一个具有足够进化深度和生态多样性的细菌门作为分析框架。系统发育基因组学分析表明,弯曲菌门最早期的进化分支由栖息于深海热液口的严格厌氧和嗜热的化能自养菌组成,该环境被认为是太古宙地球环境的模式系统,表明弯曲菌门的早期进化分支在早期地球历史中发挥了重要作用。随后的进化分支向海洋和陆地淡水系统进行生态位扩张。随着生态环境的多样化,这些谱系表现出从厌氧到微氧/好氧,从嗜热到嗜中温,以及从自养到异养的转变。其中一些谱系成为与宿主相关的共生菌或病原菌,以严格嗜中温和异养型的Campylobacter属和Helicobacter属为代表。此外,该门物种的基因组大小变化范围较窄(1.4-3.5Mb),没有发生剧烈的基因组缩减或扩张。显然,弯曲菌门是一个具有足够进化深度、分支次序有据、生态分布广泛的单系群,为研究细菌信号转导系统或信号网络的进化提供了一个很好的“生态-进化”框架(图1)。
研究人员利用弯曲菌门的“生态-进化”框架,对其门下82个物种的基因组进行了详细的比较基因组学分析。研究发现,整体上弯曲菌门物种的信号转导网络的进化模式表现出两种相反的趋势。在生态位扩张的物种中,信号网络的复杂性随着基因家族的扩张而增加,水平基因转移、基因复制、融合和裂变等新基因获得方式为基因创新和不同转导机制的整合创造了机会。相反,当细菌分支专一性适应特殊生态位时,其信号网络的复杂性随着大量基因的丢失而降低。具体到每个信号转导系统,我们发现转导机制的不同导致进化方式也有差别。主要结论包括:1.趋化系统主要通过水平基因转移获得新的整套趋化基因簇来增加复杂度,而不同趋化类别之间极少发生融合;2.双组份系统的复杂度主要取决于非经典组氨酸激酶的数量,在经典组氨酸激酶的结构域组成的基础上通过在激酶的N-端或者C-端融合REC结构域,来实现磷酸信号的多步传递、输入和输出信号的整合或分流;3.环二鸟苷酸介导的通路进化方式最多样,其合成酶和分解酶容易获得也容易丢失,因为第二信使小分子的合成和降解成本低,而且该小分子不受蛋白互作的进化约束。以上三种系统在弯曲菌门的祖先细胞中就已经进化了,后来的分支随着生态位的拓张不断增加网络复杂度;而宿主体内的共生菌和病原菌则大量丢失信号转导蛋白,只保留简单的单组份、双组份系统以及与鞭毛共进化的趋化系统,一些分支彻底丢了环二鸟苷酸介导的通路。总而言之,在弯曲菌门中,信号转导网络的大小和复杂度呈现共同增强与共同减弱的趋势,由细菌感受环境变化的需求、能量成本、遗传资源和基因组内的协同进化共同决定的。
中国科学院南海海洋研究所2022届博士莫然、2020级博士生陈园园、2020届硕士刘煜耿为该论文的共同第一作者,高贝乐研究员为通讯作者。本研究得到了国家自然科学基金、南方海洋科学与工程引进人才团队重点专项广东实验室(广州)、中国科学院战略性先导专项、中国科学院南海生态与环境工程创新研究院的资助。
相关论文信息:https://journals.asm.org/doi/10.1128/mbio.00764-22
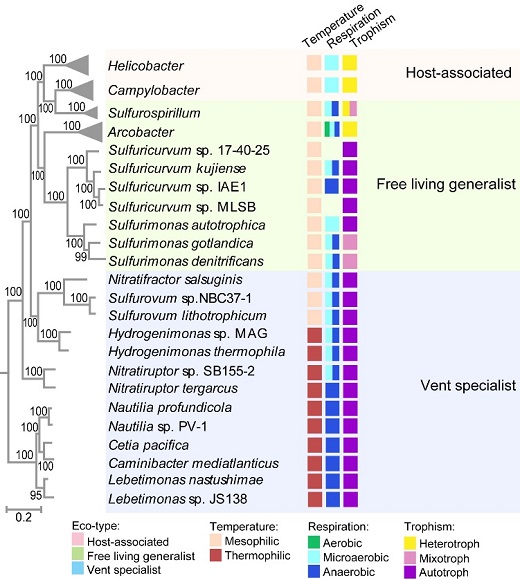
图1 弯曲菌门的物种进化树与生态生理特点
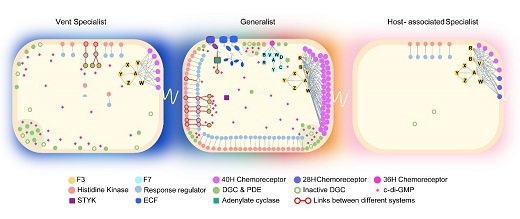
图2 弯曲菌门代表物种的信号转导网络的构成
附件下载:

 粤公网安备44011502001245号
粤公网安备44011502001245号

